

Fitomejoramiento y recursos genéticos
Caracterización morfológica y dasométrica de Mimosa trianae y Cassia moschata de la colección de AGROSAVIA
Morphological and dasometric characterization of Mimosa trianae and Cassia moschata from AGROSAVIA collection
 Sandra L. Castañeda-Garzón slcastaneda@agrosavia.co
Jessica Moreno-Barragán
Jorge H. Argüelles-Cárdenas
Hebert Camargo-Tamayo
Jhon J. Zuluaga-Peláez
Sandra L. Castañeda-Garzón slcastaneda@agrosavia.co
Jessica Moreno-Barragán
Jorge H. Argüelles-Cárdenas
Hebert Camargo-Tamayo
Jhon J. Zuluaga-Peláez
Temas Agrarios
Universidad de Córdoba, Colombia
ISSN: 0122-7610
ISSN-e: 2389-9182
Periodicidad: Semestral
vol. 26, núm. 1, 2021
Recepción: 11 Marzo 2021
Aprobación: 27 Junio 2021

Resumen: El cañafístulo (Cassia moschata Kunth) y el yopo (Mimosa trianae Benth.) son especies nativas de importancia económica en la Orinoquia Colombiana, sin embargo, se desconoce su variabilidad genética y su comportamiento en un sistema de monocultivo. En una plantación de siete años ubicada en el Centro de Investigación La Libertad (Villavicencio, Meta), se evaluó la variabilidad fenotípica de 108 árboles de M. trianae y 52 árboles de C. moschata empleando 37 descriptores cuantitativos y cualitativos de planta (copa, fuste, hoja). Se empleó un análisis de componentes principales y correspondencias múltiples, para las variables cuantitativas y cualitativas respectivamente, con el fin de reducir la dimensionalidad del conjunto de datos, complementados con un análisis de conglomerados, mediante el algoritmo de WARD. C. moschata presentó menor porte y una copa menos vigorosa, en comparación a M. trianae que se destacó por mayor número ramas, diámetro de copa, volumen de madera e incremento medio anual de la altura total y del diámetro del fuste. El análisis de conglomerados permitió clasificar los individuos de C. moschata y M. trianae en seis y cinco grupos respectivamente, reteniendo el 99.92% y el 99.58% de la variabilidad original. Para la selección de árboles maderables y su uso en sistemas agroforestales, las variables volumen de tronco, diámetro de copa, número de ramas y altura total, son las más importantes. En M. trianae el número de foliolos por hoja y el tamaño de las hojas son variables igualmente relevantes para selección de árboles de sombrío.
Palabras clave: Cañafístulo, Recursos genéticos, Variabilidad fenotípica, Yopo .
Abstract: Cañafistol (Cassia moschata Kunth) and yopo (Mimosa trianae Benth.) are important native species in the Colombian Orinoco Region, however, their behavior in a monoculture system and genetic variability are unknown. In a 7-year-old plantation located at La Libertad Research Center (Villavicencio, Meta), the phenotypic variability of 108 M. trianae trees and 52 C. moschata trees was evaluated using 37 quantitative and qualitative plant descriptors (crown, stem, leaf). Principal component analysis and multiple correspondence were used for quantitative and qualitative variables respectively, in order to reduce the dimensionality of the data, followed by cluster analysis using the WARD algorithm. C. moschata had a smaller size and a less vigorous crown, in comparison to M. trianae which stands out for its greater number of branches, crown diameter, wood volume and annual average increase of the total height and DBH. The cluster analysis allowed classifying the individuals of C. moschata and M. trianae into six and five groups retaining 99.92% and 99.58% of the original variability respectively. The variables trunk volume, crown diameter, number of branches and total height are the most important for the selection of timber trees and use in agroforestry systems. In M. trianae the number of leaflets per leaf and the size of the leaves are equally relevant variables for the selection of shade trees.
Keywords: Cañafístulo, Genetic resources, Phenotypic variability, Yopo.
INTRODUCCIÓN
En la Orinoquia Colombiana, son varias las especies forestales nativas utilizadas como leña (Acero, 2005). Entre estas se encuentran yopa (Anadenanthera peregrina (L.) Speg.), cañafístulo (Cassia moschata Kunth) y yopo (Piptadenia pteroclada Benth) (Acero, 2005; Peñuela et al., 2011; Ortega, 2015).
Mimosa trianae Benth. y C. moschata son especies nativas, de la familia Leguminosae, presentes en sistemas silvopastoriles y adaptadas a las condiciones de acidez de los suelos de la Orinoquia (Sánchez et al., 2015). En Colombia se distribuyen en las regiones biogeográficas de los Andes y la Orinoquia, aunque C. moschata se reporta igualmente en la Llanura del Caribe (Romero, 2021 y Gradstein, 2021). Su importancia económica en la Orinoquia se atribuye a que su leña se emplea como insumo en la preparación del plato típico conocido como “mamona” (carne asada) (Ortega, 2015; Calle y Murgueitio, 2020), por lo cual presenta alta demanda en el mercado interno (asaderos). El yopo utilizado en sistemas agroforestales (SAF), como sombrío por su porte y ramificación, permite en las primeras etapas de desarrollo realizar entresacas de ramas, generando un ingreso económico, mientras que las especies asociadas (p. ej. cacao) inician su etapa productiva y/o el yopo alcanza su máximo aprovechamiento.
C. moschata es medicinal, ornamental y maderable, debido a su excelente durabilidad natural, “tipifica el paisaje de la sabana como árbol de sombrío y como forraje” (Acero, 2005). M. trianae es endémica (Romero, 2021), empleada como sombrío en sistemas ganaderos o en sistemas silvopastoriles intensivos (Montagnini et al., 2015). Más que ningún otro árbol nativo de la cuenca del Orinoco, el yopo ofrece la posibilidad de lograr una integración rentable entre la ganadería y la actividad forestal en los Llanos Orientales (Calle y Murgueitio, 2020).
Con relación a la caracterización morfológica en especies forestales, Aguirre-de los Ríos y Zevallos-Pollito (2014) identificaron diferencias fenotípicas en Cordia alliodora al emplear 33 descriptores dendrológicos y 24 descriptores anatómicos de la madera. En México, la evaluación fenotípica de la especie forestal Gmelina arborea, incluyó las variables altura del árbol (HT), diámetro a la altura del pecho (DAP), volumen del tronco, rectitud del fuste (RFU), diámetro de la copa (D_COPA), diámetro de la primera rama viva, ángulo de inserción de ramas (AIR) y densidad de la madera (Balcorta y Hernández, 2004). Para Juglans pyriformis, las variables empleadas fueron DAP, HT, forma de fuste, altura de bifurcación, dominancia del eje principal, AIR, forma y diámetro de copa (Acosta-Hernández et al., 2011). Por otra parte, para la selección de árboles plus de Alnus acuminata, Cedrela sp. y Nectandra sp. en Perú (Oliva y Remachi, 2017), se utilizaron las variables DAP, HT y RFU. Para la estimación de parámetros genéticos de ensayos de progenie de Pinus patula (Escobar-Sandoval et al., 2018) se evaluaron las variables HT, DAP y AIR, entre otras asociadas a calidad de madera y ramificación. La morfometría de copa y rasgos foliares se ha evaluado en Araucaria angustifolia (Hess et al., 2016), Abatia parviflora, Escallonia paniculata y Quercus humboldtii (Melo-Cruz et al., 2012), Caesalpinia gaumeri y Gymnopodium floribundum (Interián-Ku et al., 2009).
El proyecto de investigación “Evaluación de especies arbustivas y arbóreas para uso múltiple en diferentes agroecosistemas de Colombia”, ejecutado por parte de la Corporación Colombiana de Investigación Agropecuaria-AGROSAVIA, incluyó entre otros aspectos, el uso de descriptores morfológicos y dasométricos en las especies M. trianae y C. moschata, con el objetivo de evaluar la variabilidad fenotípica de la colección de trabajo, como insumo para la identificación de individuos sobresalientes para sistemas agroforestales, producción de madera u otros usos.
MATERIALES Y MÉTODOS
Los lotes correspondientes a las colecciones de trabajo de las especies forestales del Centro de Investigación de AGROSAVIA La Libertad, en Villavicencio-Meta (4°03´50” N, 7°32´53” W a 342 msnm), están ubicados en el piedemonte llanero, en terreno llano, zona de planicie, entorno de llanura anivelada y pendiente plana. La conforman 108 árboles de 7 años de edad de M. trianae y 52 árboles de C. moschata, sembrados en monocultivo a una distancia de 5x5 m y 4x4 m respectivamente, para facilitar la evaluación de la producción de leña y evitar la competencia entre copas de los árboles. El material vegetal propagado por semilla se adquirió en vivero comercial, sin distinción de procedencia o progenie.
Para la caracterización morfológica de la totalidad de individuos de la colección, se evaluaron 37 descriptores (22 de árbol y 15 de hoja) (Tabla1). El color se registró empleando la tabla de tejidos (Munsell) y para la medición de las variables cuantitativas, se utilizó pértiga telescópica (Hastings M-40), cinta diamétrica (Lufkin), pie de rey digital (Mitutoyo) y flexómetro (Stanley). Para la evaluación de descriptores foliares se recolectaron cinco hojas maduras por árbol, completas y sin problemas fitosanitarios; muestreando cinco foliolos (por hoja) ubicados a los dos costados en la parte media.
Para conocer el desarrollo de las especies, durante siete años y con periodicidad semestral se evaluaron las variables dasométricas altura total y diámetro a la altura del pecho (DAP) medido a los 1,3 m sobre el nivel del suelo (a partir de los 29 meses una vez los individuos superaron esta altura, los registros anteriores se tomaron en la base del tallo). Posteriormente se calculó el DAP medio y la altura total media, el porcentaje de sobrevivencia y el incremento medio anual (IMA) para DAP y HT, el cual se determinó teniendo en cuenta la diferencia entre la última y la primera medición, dividido por el tiempo trascurrido entre las dos mediciones (López et al., 2015).
| Descriptor | Sigla | Autor |
| Altura del árbol (m) | HT | 7, 8 |
| Altura de copa (m) | HC | 7 |
| Longitud de la copa (sentido NS) - m | LCNS | 7, 8 |
| Longitud de la copa (sentido EW) - m | LCEW | 7, 8 |
| Diámetro a la altura del pecho (cm) | DAP | 8 |
| Volumen de tronco (m³) | VOL_TRONCO | 5 |
| Diámetro de copa (m) | D_COPA | 8* |
| Número de ramas principales | NRAM | 6* |
| Forma del árbol | FARB | 8 |
| Hábito de crecimiento del árbol | HABARB | 8 |
| Forma del fuste | FFU | 6 |
| Rectitud del fuste | RFU | 1* |
| Ramificación del árbol | RAM | 3 |
| Altura de ramificación | HRAM | 1* |
| Altura de bifurcación | HB | 1* |
| Dominancia en el eje principal | DEP | 1* |
| Ángulo de inserción de las ramas | AIR | 1, 9 |
| Forma de la copa (perfil vertical) | FCV | 7* |
| Forma de la copa (perfil horizontal) | FCH | 4* |
| Densidad de la copa | DNC | 10 |
| Tipo de corteza | CORT | 3* |
| Color del tronco | CT | 9* |
| Número de foliolos por hoja | NFH | 3* |
| Forma de la hoja | FH | 7, 8* |
| Borde de la hoja | BORH | 7, 8* |
| Ápice de la hoja | APH | 7, 8* |
| Base de la hoja | BH | 7, 8* |
| Longitud del peciolo (mm) | LPEC | 8* |
| Longitud del peciolo principal (mm) | LPECppal | 2* |
| Descriptor | Sigla | Autor |
| Longitud del raquis (mm) | LR | 2* |
| Grosor del peciolo (mm) | GPEC | 2 |
| Longitud de la hoja (mm) | LH | 2 |
| Anchura de la hoja (mm) | ANH | 2 |
| Longitud del foliolo (mm) | LHF | 2* |
| Anchura del foliolo (mm) | ANHF | 2* |
| Color de las hojas maduras en el haz | CHMHAZ | 8* |
| Color de las hojas maduras en el envés | CHMENV | 8* |
Para reducir la dimensionalidad del conjunto de datos, se realizó un análisis de correspondencia múltiple (ACM) para las variables cualitativas, mediante el procedimiento CORRESP de SAS Enterprise Guide V.8.3 (2019-2020) y un análisis de componentes principales (ACP) para las variables cuantitativas, utilizando el procedimiento PRINCOMP del citado software. Posteriormente y con base en las dimensiones seleccionadas en los dos procedimientos estadísticos anteriores, se realizó un análisis de conglomerados (AC), con el propósito de clasificar los individuos en grupos homogéneos, mediante el uso del procedimiento CLUSTER (algoritmo de Ward, distancia euclidiana) del mencionado software. Para cada uno de los grupos conformados, se generaron estadísticas básicas, que permitieron caracterizar dichos grupos.
RESULTADOS Y DISCUSIÓN
En la colección de trabajo de C. moschata y M. trianae del Centro de Investigación La Libertad, se observó variación en los rasgos cuantitativos de planta. La tabla 2 se construyó a partir de las frecuencias de individuos por descriptor, como apoyo a la descripción general. De acuerdo con las categorías referidas por Quiceno et al. (2016) para calificar la forma de la copa, el descriptor FCH reveló que C. moschata exhibe una copa muy pobre al estar conformada principalmente por rebrotes, en contraste con una copa buena/vigorosa y tolerante en M. trianae por su forma predominantemente circular irregular y de medio círculo (Tabla 2). Los árboles de M. trianae presentaron mayor NRAM (Tabla 3) ramificándose en el tercio inferior, característica que la hace una especie apetecida en el mercado para el consumo de leña.
En Acacia mangium Willd., la presencia y la altura de bifurcación son algunas de las variables evaluadas para determinar la calidad de madera. Pavlotzky-Blank y Murillo-Gamboa (2013) identificaron que “a mayor altura de bifurcación mejor es la calidad del árbol y a mayor presencia de bifurcación peor es la calidad”. Aunque C. moschata y M. trianae se utilizan en la Orinoquia para producción de leña (Acero, 2005; Calle y Murgueitio, 2020), la selección de árboles para la producción de biomasa y madera podría llevarse a cabo integrando variables del fuste, arquitectura de la copa y poder calorífico. Nutto et al. (2001) mencionan que variables como la superficie, el diámetro y la longitud del dosel están directamente relacionados con el crecimiento y la producción de materia seca del árbol. No obstante, para evaluar la arquitectura del dosel se requeriría determinar la cantidad y distribución espacial de las ramas, hojas y frutos, e incluso el índice de área foliar (Castaño et al., 2013).
| Descriptor | C.moschata | M. trianae |
| FARB | Extendido (90,38%) | Extendido (95,83%) |
| HABARB | Extendido (100%) | Extendido (94,17%) |
| FFU | Cónico (96,15%) | Cilíndrico (99,17%) |
| RFU | Algunas curvaturas en más de un plano (44,23%) | Torcido (90,83%) |
| RAM | Acodillada (98,08%) | Acodillada (65,83%) |
| HRAM | En el tercio medio (55,77%) | En el tercio inferior (98,33%) |
| NRAM | Dos (57,69%) | Tres (42,50%) |
| HB | En el tercio inferior (63,46%) | No bifurcado (65%) |
| DEP | Parcial del eje inicial sobre las ramas laterales (100%) | Parcial del eje inicial sobre las ramas laterales (79,17%) |
| AIR | ≥ 31° y 60° (94,23%) | ≥ 61° - 90° (90,83%) |
| FCV | Irregular (84,62%) | Extendida (95%) |
| FCH | Principalmente rebrotes (40,38%) | Circular irregular (27.50%), medio círculo (21,67%) |
| DNC | Densa (53,85%) | Intermedia (34,17%) |
| TCORT | Surcada vertical (100%) | Papelosa (99,17%) |
| CT | 2.5GY6/2; 2.5GY7/2 (30,77% y 19,23%) | 5Y8/2; 5YR5/4; 5YR4/2 (66,67%) |
| FH | Oblonga (92,71%) | Oblonga (100%) |
| BORH | Entero (100%) | Entero (100%) |
| APH | Obtuso (74,75%) | Mucronado (69,31%) |
| BH | Truncada (73,80%) | Truncada (59,64% |
| CHMHAZ | 5GY4/6 (76,08%) | 5GY (5/4, 4/4, 5/6, 5/8, 5/10, 4/6, 4/8, 6/6, 7/8, 6/8 (72,09%) |
| CHMNENV | 5GY6/8 (83,14%) | 5GY7/4 (71,16%) |
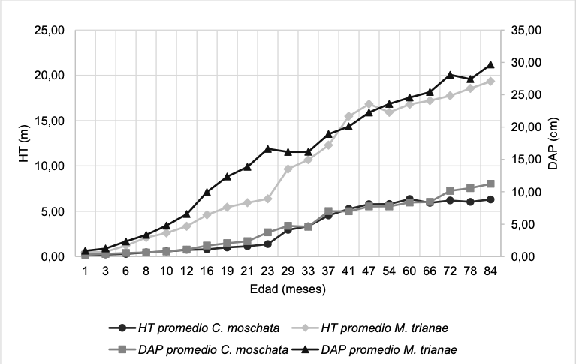
En la colección de trabajo, las variables VOL_ TRONCO y D_COPA revelaron mayores valores en M. trianae, aspecto que podría atribuirse al rápido crecimiento (Tabla 3). El diámetro de la copa refleja la dimensión del aparato fotosintético del árbol, aspecto directamente relacionado con su capacidad de crecimiento (Arias, 2005). Durante los primeros siete años, el crecimiento de C. moschata y M. trianae fue diferente en términos de la HT y el DAP (Figura 1). C. moschata presentó una menor tasa de crecimiento (IMAHT: 0,9 m e IMADAP: 1,6 cm), en comparación a M. trianae (IMAHT: 1,86 m e IMADAP: 2,63 cm). En el municipio de Acacias (Meta), Gutiérrez (2014) observó en plantas M. trianae de 1,3 años de edad, IMA en altura de 1,76 m en monocultivo y de 1,72 m en arreglo múltiple; superando lo obtenido en C. moschata (IMA de 1,04 m y 0,96 m) en plantas de la misma edad y sistema de cultivo. Algunas de las características observadas en los individuos de C. moschata (Tabla 2 y Tabla 3) concuerdan con la descripción de “árbol mediano, de copa irregular y amplia, follaje caducifolio” (Rojas-Rodríguez y Torres- Córdoba, 2008), copa ancha y extendida (CAR, 2012), corteza externa fisurada y de color pardo oscuro (Ariza et al., 2016).
| Descriptor | Media (C. moschata) | IC < (C. moschata) | IC > (C. moschata) | Media (M.trianae) | IC < (M. trianae) | IC >(M. trianae) |
| HT (m) | 6,3 | 5,89 | 6,71 | 10,65 | 10,29 | 11,01 |
| HC (m) | 1,37 | 1,26 | 1,48 | 6,43 | 6,14 | 6,71 |
| LCNS (m) | 6,17 | 5,72 | 6,63 | 7,73 | 7,36 | 8,1 |
| LCEW (m) | 6,13 | 5,64 | 6,62 | 7,66 | 7,28 | 8,04 |
| DAP (m) | 11,22 | 10,06 | 12,4 | 16,97 | 16,13 | 17,82 |
| VOL_TRONCO (m³) | 0,03 | 0,02 | 0,04 | 0,2 | 0,18 | 0,22 |
| D_COPA (m) | 6,15 | 5,73 | 6,57 | 7,7 | 7,37 | 8,02 |
| NRAM | 1,94 | 1,75 | 2,14 | 3,05 | 2,82 | 3,27 |
| NFH | 29,68 | 28,81 | 30,55 | 26,11 | 25,59 | 26,62 |
| LPEC (mm) | 156,76 | 150,29 | 163,24 | 82,82 | 81,22 | 84,43 |
| LPECppal (mm) | 18,48 | 17,63 | 19,32 | NA | NA | NA |
| LR (mm) | 1,55 | 1,48 | 1,62 | 10,58 | 10,36 | 10,8 |
| GPEC (mm) | 1,28 | 1,25 | 1,31 | 1,66 | 1,62 | 1,71 |
| LH (mm) | 43,89 | 42,1 | 45,67 | 150,7 | 147,99 | 153,41 |
| ANH (mm) | 14,44 | 14,02 | 14,87 | 88,74 | 86,86 | 90,63 |
| LHF (mm) | NA | NA | NA | 54,96 | 53,93 | 55,99 |
| ANHF (mm) | NA | NA | NA | 7,6 | 7,4 | 7,81 |
“Presenta de 10 a 20 pares de hojuelas de 4 a 5 cm de largo y 1,5 cm de ancho” (Rojas- Rodríguez y Torres-Córdoba, 2015), “foliolos oblongo-obovolados, ápice redondeado mucronado, base redondeada y margen entero” (Ariza et al., 2016). Así mismo, algunos rasgos de M. trianae son semejantes a la descripción “árbol de copa expandida y rala, corteza rugosa y de color café o gris; las hojas de 12 a 30 cm de largo son bipinnadas, alternas y tienen 10 a 20 pares de foliolos” (Giraldo, 2020).
Producto del ACM se seleccionaron para C. moschata y M. trianae las primeras cinco y tres dimensiones, que acumularon el 84,75% y el 86,89% de la variabilidad contenida en las variables cualitativas originales respectivamente. El ACP basado en la matriz de correlaciones, permitió seleccionar para C. moschata y M. trianae los primeros cinco componentes principales (CP), que acumularon el 85,04% y 80,80% de la variabilidad contenida en las variables cuantitativas originales respectivamente. A partir de las dimensiones o los componentes seleccionados en el ACM y en el ACP, se generó un conjunto de datos integrando estos dos tipos de variables sintéticas, para realizar el AC que permitió clasificar los individuos de C. moschata y M. trianae en seis y cinco grupos, reteniendo el 99,92% y el 99,58% de la variabilidad contenida en las anteriores variables, respectivamente (Figura 2 y Figura 3). En M. trianae, las variables de mayor aporte en la discriminación de los árboles fueron D_COPA, VOL_TRONCO, NRAM y HT; mientras que en C. moschata fueron D_COPA, LR, NRAM, LPECppal y LCNS.
En C. moschata se identificaron individuos con potencial de uso maderable en los grupos 2 (25%) y 3 (19,23%) considerando el mayor volumen de madera por árbol individual (0,08 a 0,14 m3) según lo expresado por Vallejos et al. (2010). Estos árboles podrían ser útiles como sombrío, por presentar mayor D_COPA (7,61 a 10,30 m) y NRAM superior a 3. El hábito de fuste múltiple es un atributo que le otorga la posibilidad de uso para producción de leña y sombrío (Raintree, 1996).
En el grupo 1 (15,38%) se observaron árboles con potencial de uso para producción de leña en monocultivo, al presentar HT intermedia (5,78 a 7,57 m), HC intermedia a mayor (1,04 a 2,53 m), mayor VOL_TRONCO (0,08 a 0,14 m3), DAP (10,77 a 15,16 cm) y D_COPA (5,81 a 7,60 m) intermedio; NRAM intermedio (3 ramas), RFU ligeramente torcido, HRAM en el tercio medio, HB en el tercio inferior, FCH menos de un tercio de círculo, DNC intermedio y mayor NFH (31 a 35).
Aunque los individuos de C. moschata del grupo 4 (17,31%) presentaron HT intermedia (5,78 a 7,57 m), NRAM mayor a 3, VOL_ TRONCO mayor (0,08 a 0,14 m3), HRAM en el tercio medio y hojas de tamaño intermedio (LH 43,17 a 50,15 mm y ANH 13,81 a 15,80 mm), la forma de la copa principalmente de rebrotes es catalogada como muy pobre para ofrecer sombra (Quiceno et al., 2016). Esta última característica es desfavorable para sombrío en SAF con cacao, ya que esta especie requiere entre 50% y 70% de sombra en los primeros años de desarrollo, para que la planta logre mayores tasas fotosintéticas (Agudelo- Castañeda et al., 2018; Ramírez et al., 2020). Aunque en los grupos 5 (9,62%) y 6 (13,46%) de C. moschata se identificaron árboles de menor tamaño de copa (3,10 a 5,8 m), el grupo 6 se caracterizó por menor HT (3,07 a 5,77 m) y menor HC (0,13 a 1,03 m), característica que podría facilitar la cosecha de ramas según lo expuesto por Raintree (1996), pero no sería útil para ofrecer sombra. A pesar de que el grupo 5 tuvo individuos con mayor NRAM y copa exclusivamente densa, presenta menor D_ COPA y HT, características desfavorables para sombrío en la etapa juvenil de otras especies establecidas en SAF.


Por otra parte, en la colección de 108 árboles de M. trianae el AC permitió clasificarlos en cinco grupos (Figura 3), se identificaron individuos con potencial maderable en el grupo 2 (14,81%) por el mayor VOL_TRONCO (0,31 a 0,48 m3). El mayor D_COPA (8,41 a 12,90 m), NFH (27 a 31) y tamaño de hojas (ANH: 94,72 a 115,71 mm; LHF: 57,81 a 68,30 mm) y copa de forma circular irregular, le atribuyen potencial de uso como sombrío. Los foliolos pequeños de algunas especies leguminosas proporcionan “una sombra rala” y por lo tanto, menor frecuencia de podas (Somarriba, 1996), aspecto que favorecería el manejo de la sombra en el cultivo de cacao.
En los grupos 1 (37,96%) y 3 (26,85%) se observaron árboles de porte medio con respecto a HT (9,16 a 12,75 m) y HC (4,94 a 7,13 m), copa mediana (D_COPA: 5,41 a 8,40 m), menor VOL_TRONCO (0,00 a 0,18 m3) y NRAM (1 a 3). No obstante, en el primer grupo se observaron árboles con FCH predominantemente circular irregular, DNC intermedio, mayor NFH (17 a 22) y menor tamaño de hojas (LR, LH, ANH, LHF, ANHF). El tercer grupo se destacó por la presencia de árboles de copa pobre, conformados por pocas ramas (Quiceno et al., 2016). Aunque estos grupos carecieron de características deseables para sombrío, su HT, HC y NRAM podrían conferirle uso potencial maderable, esto, teniendo en cuenta lo expuesto por Palencia et al. (2006), al indicar que entre los criterios de selección de especies forestales para uso en SAF con cacao, se deben considerar especies de rápido crecimiento, “para lograr que el tiempo estimado del retorno de la inversión sea atractivo para el inversionista”.
Los árboles del grupo 4 (14,81%) podrían ser igualmente útiles como sombrío y producción de leña al presentar hábito de fuste múltiple debido al mayor NRAM (6 a 8), así como HT y HC intermedia (9,16 a 12,75 m y 4,94 a 7,13 m respectivamente) que facilitaría la cosecha de ramas; estas características son resaltadas por Raintree (1996) para las especies de uso múltiple. Los grupos 4 y 5 (5,56%) exhibieron copa de forma circular irregular catalogada como vigorosa por Quiceno et al. (2016) e igualmente útil para sombrío.
CONCLUSIONES
El método de evaluación de descriptores de planta proporcionó información útil para la evaluación de la variabilidad fenotípica de la colección de trabajo de C. moschata y M. trianae, así como la identificación preliminar de árboles con potencial de uso maderable y asocio con SAF.
En C. moschata los conglomerados 2 y 3 agrupan árboles con potencial de uso maderable y sombrío, atribuido al mayor volumen del tronco, diámetro de copa y número de ramas. Los árboles del grupo 6 son candidatos de selección para cosecha de leña, debido a la menor altura total y altura de copa. Para la producción de leña en monocultivo, las variables útiles en la selección de árboles en el grupo 1 son altura total, altura de copa, volumen de tronco, DAP, número de ramas y altura de ramificación.
Para la selección de árboles de M. trianae con potencial de uso maderable se podrán contemplar individuos de los grupos 1 y 3 que se destacan por mayor altura total, altura de copa y número de ramas principales. Y para uso como sombrío en SAF, la selección de árboles se podría realizar en el grupo 2 que presenta mayor volumen del tronco, diámetro de copa, tamaño de hoja y número de foliolos por hoja.
Agradecimientos
Los autores desean agradecer a AGROSAVIA adscrita al Ministerio de Agricultura y Desarrollo Rural de Colombia (MADR) por financiar este estudio. A Ángela Villa, Arnoldo Muñoz, Oscar Triana y Jennifer Rodríguez, por su apoyo en la recolección de material vegetal y procesamiento.
REFERENCIAS
Acosta-Hernández, C., Luna-Rodríguez, M., Noa-Carrazana, J., Galindo-González, J., Vázquez-Torres, S., Morales-Romero, Z. e Iglesias-Andreu, L. 2011. Caracterización morfológica y dasométrica de la especie amenazada Juglans pyriformis Liebm. Revista Chapingo Serie Ciencias Forestales y Del Ambiente XVII (1):59–67.
Agudelo-Castañeda, G., Cadena-Torres, J., Almanza-Merchán, P., Pinzón-Sandoval, E. 2018. Desempeño fisiológico de nueve genotipos de cacao (Theobroma cacao L.) bajo la sombra de tres especies forestales en Santander, Colombia. Revista Colombiana de Ciencias Hortícolas 12(1):223-232.
Aguirre-de los Ríos, F., Zevallos-Pollito, P. 2014. Caracterización fenotípica, dendrológica y anatómica de los tipos morfológicos del Cordia alliodora (R. & P.) Oken procedentes de plantaciones realizadas en Jaén, Cajamarca-Perú. El CEPROSIMAD 2(2): 7-20.
Arias, D. 2005. Morfometría del árbol en plantaciones forestales tropicales. Kurú: Revista Forestal 2(5):1-13.
Bioversity International, CHERLA. 2008. Descriptores para chirimoyo (Annona cherimola Mill.). Bioversity International. https://www.bioversityinternational.org/fileadmin/_migrated/uploads/tx_news/ Chirimoyo_1295.pdf
Calle, Z., Murgueitio E. 2020. Árboles nativos para predios ganaderos. Especies focales del Proyecto Ganadería Colombiana Sostenible. CIPAV, Cali Colombia. https://elti.yale.edu/sites/default/files/ rsource_files/arboles_nativos_para_ predios_ganaderos.pdf
Castaño, A., Chica, H., Obando, D., Riaño, N. 2013. Arquitectura del dosel e interceptación de la radiación solar por tres especies forestales nativas en Colombia. Cenicafé 64 (1):19-30.
Giraldo, A. 2020. Fenología, crecimiento y diversidad genética de Mimosa trianae Benth en el Piedemonte Orinocense y el Valle del Cauca, Colombia. Tesis de maestría en Ciencias Biológicas con énfasis en recursos fitogenéticos. Universidad Nacional de Colombia, Palmira.
Gradstein, S. 2021. Cassia moschata Kunth. En Bernal, R., S.R. Gradstein & M. Celis (eds.). 2015. Catálogo de plantas y líquenes de Colombia. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. http://catalogoplantasdecolombia.unal.edu.co/es/resultados/especie/Cassia%20moschata/
Gutiérrez, E., Moreno R., Villota, N. 2013. Guía de cubicación de la madera. Corporación Autónoma Regional del Risaralda – CARDER. Pereira, Colombia, p12.
Gutiérrez, A. 2014. Aportes a la rehabilitación ecosistémica de áreas riparias con énfasis en calidad de suelos y producción dendroenergética por medio de arreglos agroforestales en zonas ganaderas del piedemonte llanero. Tesis de maestría en Agroforestería Tropical. Universidad de Ciencias Ambientales y Aplicadas U.D.C.A, Bogotá.
Hess, A., Loiola. T., Arruda de Souza, I., Nascimento, B. 2016. Morfometría de la copa de Araucaria angustifolia en sitios naturales en el sur de Brasil. Bosque 37(3):603-611.
López, L., Villalba, R. 2015. Criterios de gestión forestal para 12 especies de los Bosques Nativos Tropicales de Bolivia a través de métodos dendrocronológicos. Ecosistemas 24(2):24-29.
Instituto Internacional de Recursos Fitogenéticos - IPGRI. 1996. Descriptores del café (Coffea spp. y Psilanthus spp.). Roma. https://www.bioversityinternational.org/ fileadmin/_migrated/uploads/tx_news/ Descriptores_del_caf%C3%A9 Coffea_ spp._y_Psilanthus_spp. 487.pdf
Instituto Internacional de Recursos Fitogenéticos - IPGRI. 2000. Descriptors for Jackfruit (Artocarpus heterophyllus). Roma.
Interián-ku, V., Valdez-Hernández, J., García- Moya, E., Romero-Manzanares, A., Borja-De-La-Rosa, M., Vaquera-Huerta, H. 2009. Arquitectura y morfometría de dos especies arbóreas en una selva baja caducifolia del sur de Yucatán, México. Boletín de la Sociedad Botánica de México 85:17-29.
Kehlenbeck K., S. Padulosi, A. Alercia. 2015. Descriptors for Baobab (Adansonia digitata L.). Bioversity International. Roma. http://www.bioversityinternational.org/ fileadmin/user_upload/online_library/ publications/pdfs/Descriptors_for_ Baobab__Adansonia_digitata_L.__1859. pdf
Melo-Cruz, O., Rodríguez-Santos, N. y Rojas- Ramírez, F. 2012. Patrones de arquitectura foliar asociados al crecimiento funcional de cinco especies leñosas nativas de la cordillera oriental utilizadas en restauración ecológica en la Sabana De Bogotá. Colombia Forestal 15(1):119- 130.
Montagnini, F., Somarriba, E., Murgueitio, E., Fassola, H., Eibl, B. 2015. Sistemas Agroforestales. Funciones Productivas, Socioeconómicas y Ambientales. Serie técnica. Informe técnico 402. CATIE, Turrialba, Costa Rica. Editorial CIPAV. Cali, Colombia.
Nutto, L. Tonini, H. Borsoi, G., Moskovich, F., Spathelf, P. 2001. Utilização dos parâmetros da copa para avaliar o espaço vital em povoamentos de Pinus elliottii Engelm. Boletim de Pesquisa Florestal 42:123-138.
Ortega, E. 2015. Usos tradicionales de las plantas de la Orinoquia colombiana. UG-Ciencia 21:16-28.
Ortiz, E., Acosta, C., Linares, P., Morales, Z., Rebolledo, V. 2017. Selección de árboles semilleros de Juglans pyriformis Liebm. en poblaciones naturales de Coatepec y Coacoatzintla, Veracruz. Revista Mexicana de Ciencias Forestales 7(38):43-58.
Palencia, E., Gómez, R., Martín, J., Guiza, O. 2006. Especies forestales para uso en sistemas agroforestales con cacao. Produmedios. Bogotá. 12p. http://cadenacacaoca.info/CDOC- Deployment/documentos/Especies_ forestales_para_uso_en_sistemas_ agroforestales_con_cacao.pdf
Pavlotzky-Blank, B., Murillo-Gamboa, O. 2013. Ganancia genética esperada en Acacia mangium en San Carlos, Zona Norte de Costa Rica. Revista Forestal Mesoamericana Kurú 10(24):14-25.
Peñuela, L., Castro, F., Ocampo-Peñuela, N. 2011. Las reservas naturales del Nodo Orinoquia en su rol de conservación de la biodiversidad. Fundación horizonte verde y Resnatur, Bogotá, p34.
Quiceno, N., Tangarife, G., Álvarez, R. 2016. Estimación del contenido de biomasa, fijación de carbono y servicios ambientales, en un área de bosque primario en el Resguardo Indígena Piapoco Chigüiro-Chátare de Barrancominas, departamento del Guainía (Colombia). Revista Luna Azul 43:171-202.
Raintree, J. 1996. Características socioeconómicas de los árboles y de las prácticas de plantación forestal. Capítulo 2 Los atributos socioeconómicos de los árboles. Organización de las Naciones Unidas para la Agricultura y la Alimentación –FAO. Roma. http://www.fao.org/3/u4377s/U4377S00. htm#TOC
Ramírez, L., Abaunza, C., Rodríguez, L., Varón, E., Barragán, E., Rojas, J. 2020. Modelo productivo para el cultivo de cacao (Theobroma cacao) para el departamento del Huila. Corporación Colombiana de Investigación Agropecuaria (AGROSAVIA). Mosquera, Colombia. https://doi.org/10.21930/agrosavia. model.7403558
Rojas-Rodríguez, F., Torres-Córdoba, G. 2008. Árboles del Valle Central de Costa Rica: reproducción Caoba. Kurú: Revista Forestal 5(14):1-3.
Romero, C. 2021. Mimosa trianae Benth. En Bernal, R., S.R. Gradstein & M. Celis (eds.). 2015. Catálogo de plantas y líquenes de Colombia. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. http://catalogoplantasdecolombia. unal.edu.co/es/resultados/especie/ Mimosa%20trianae/
Saavedra-Romero, L. de L., Alvarado-Rosales, D., Hernández-de la Rosa, P., Martínez- Trinidad, T., Mora-Aguilera, G., Villa- Castillo, J. 2016. Condición de copa, indicador de salud en árboles urbanos del Bosque San Juan de Aragón, Ciudad de México. Madera Bosques 22(2):15-27.
Sánchez, V., Delreal, G., Plazas, C., Pérez, G. 2015. Factibilidad económica de la asociación maíz-pasto en el establecimiento de un sistema silvopastoril en el piedemonte llanero de Colombia. Pastos y Forrajes 38(1): 73-79.
Somarriba, E., Beer, J., Bonnemann, A. 1996. Árboles leguminosos y maderables como sombra para cacao: el concepto. Serie Técnica No. 274. CATIE. Turrialba, Costa Rica. p19.
Thirakul, S. 1998. Manual de dendrología para 146 especies forestales del Litoral Atlántico de Honduras. Corporación Hondureña de Desarrollo Forestal, AFE/ COHDEFOR. Atlántida, Honduras, p10.
Información adicional
Conflicto de intereses: Los autores declaran que es un trabajo original y no existió conflicto de intereses de
ningún tipo en la elaboración y publicación del manuscrito.

