

Producción de cultivos y poscosecha
Correlaciones y análisis de sendero entre características del fruto y la semilla de Benincasa hispida [(Thunb.) Cogn.]
Correlations and path analysis between fruit characters and seeds of Benincasa hispida [(Thunb.) Cogn].
 Miguel M. Espitia-Camacho mmespitia@correo.unicordoba.edu.co
Hermes Araméndiz-Tatis
Carlos E. Cardona-Ayala
Miguel M. Espitia-Camacho mmespitia@correo.unicordoba.edu.co
Hermes Araméndiz-Tatis
Carlos E. Cardona-Ayala
Temas Agrarios
Universidad de Córdoba, Colombia
ISSN: 0122-7610
ISSN-e: 2389-9182
Periodicidad: Semestral
vol. 26, núm. 1, 2021
Recepción: 21 Marzo 2021
Aprobación: 27 Junio 2021

Resumen: Las correlaciones y el análisis de sendero son estrategias importantes para mejorar la selección de características agronómicas de interés comercial en diversos cultivos. El objetivo de este trabajo fue estimar las correlaciones entre 14 características biométricas del fruto y la semilla, y el análisis de sendero para el peso del fruto en B. hispida. Se evaluaron 10 familias de hermanos medios, en un diseño completamente aleatorizado con cinco repeticiones. El peso del fruto mostró correlaciones fenotípicas significativas directas con grosor de la pulpa, largo del fruto, ancho del fruto, número de semillas, cavidad semilla y peso de semillas, cuyos coeficientes oscilaron entre 0,64* y 0,91**, respectivamente. El largo de semilla presentó asociación directa significativa con volumen de una semilla y el peso de una semilla, con registros de 0,66* y 0,83**, respectivamente. El ancho de semilla y el volumen de una semilla acusaron correlación de 0,88**, mientras que la densidad de una semilla presentó correlación inversa significativa con el ancho de semilla y el volumen de una semilla, con registros de -0,73* y -0,89**. Los efectos indirectos explicaron principalmente la correlación significativa del peso del fruto con las variables ancho del fruto, cavidad semilla, peso de semillas y número de semillas. El largo del fruto presentó los mayores efectos directos e indirectos sobre el peso del fruto. El peso del fruto y largo del fruto, pueden ser usados en la selección de B. hispida para la obtención de genotipos con mayor peso y número de semillas por fruto.
Palabras clave: Asociación, Calabacín chino, Cucurbitaceae, Efectos directos e indirectos, Mejoramiento genético.
Abstract: Correlations and path analysis are important strategies to improve the selection of agronomic traits of commercial interest in various crops. The objective of this work was to estimate the correlations between 14 biometric characteristics of the fruit and the seed, and the path analysis for the weight of the fruit in B. hispida. 10 families of half siblings were evaluated, in a completely randomized design with five repetitions. Fruit weight showed direct significant phenotypic correlations with pulp thickness, fruit length, fruit width, number of seeds, seed cavity and seed weight, whose coefficients ranged between 0.64 * and 0.91 **, respectively. Seed length showed a significant direct association with the volume of a seed and the weight of a seed, with records of 0.66 * and 0.83 **, respectively. The width of the seed and the volume of a seed showed a correlation of 0.88 **, while the density of a seed presented a significant inverse correlation with the width of the seed and the volume of a seed, with records of -0.73 * and -0.89 **. Indirect effects mainly explained the significant correlation of fruit weight with the variables fruit width, seed cavity, seed weight and number of seeds. The length of the fruit presented the greatest direct and indirect effects on the weight of the fruit. The weight of the fruit and the length of the fruit can be used in the selection of B. hispida to obtain genotypes with greater weight and number of seeds per fruit.
Keywords: Association, Chinese zucchini, Cucurbitaceae, Direct and indirect effects, Genetic improvement.
La especie Benincasa. hispida [(Thunb.) Cogn.], es una Cucurbitaceae, alógama que se cultiva principalmente desde el Sur y Este de Asia (Ekeke et al., 2019; Chomicki et al., 2020); valorada principalmente por su larga vida útil y buen margen de valor agregado. Los frutos se consumen de diferentes formas (Pradhan et al., 2020a), por lo que es de gran importancia en la alimentación, nutrición, biomedicina, medicina tradicional e industria farmacológica, en las zonas productoras (Gupta et al., 2019a, 2019b; Chomicki et al., 2020).
En los programas de mejoramiento genético es de gran interés conocer la magnitud, sentido y naturaleza de las correlaciones (Nansikombi et al., 2019), al igual que el análisis de sendero, el cual permite desdoblar el coeficiente de correlación (fenotípico, genético o ambiental), en los efectos directos e indirectos de varios caracteres sobre una variable básica (Pradhan et al., 2020b). De esta manera, se aumenta la eficacia de la selección entre los caracteres de interés biológico y agronómico para obtener nuevos cultivares con varios caracteres mejorados simultáneamente (Tratenmuller et al., 2019).
El conocimiento e interpretación de las correlaciones y el análisis de sendero de las características biométricas del fruto y la semillas, es relevante para definir la estrategia más eficaz en el mejoramiento genético y estimar el progreso genético para tales caracteres; la selección basada en los efectos directos e indirectos es mucho más eficaz y útil que la selección por rendimiento per se (Tratenmuller et al., 2019; Pradhan et al., 2020b).
Estudios sobre correlaciones y/o análisis de sendero entre el rendimiento, características biométricas del fruto y semilla, en B. hispida han sido previamente reportados (Nagaraju et al., 2016; Pradhan et al., 2020b), con valores diferenciales para la asociación entre los caracteres, en cuanto a su magnitud y sentido al igual que su descomposición en efectos directos e indirectos, mediante el análisis de sendero.
El peso y el largo del fruto pueden ser usados como criterios de selección, en el mejoramiento genético de B. hispida para la selección y obtención de genotipos con mayor producción, cavidad semilla, peso de semillas y número de semillas por fruto. Esta situación es favorable para el fitomejorador y el productor, por su interés en la selección de genotipos de alta producción de frutos y semillas (Nagaraju et al., 2016; Pradhan et al., 2020b). No obstante, se debe tener en cuenta la relación entre el peso por fruto y el grosor de la pulpa, cuando el objetivo del mejoramiento es aumentar la eficiencia fisiológica en la producción de pulpa, que es la parte de mayor interés del fruto junto con las semillas (Pradhan et al., 2020b).
En Colombia, B. hispida ha sido poco investigada, no existen reportes sobre correlaciones y análisis de sendero para el peso del fruto y sus componentes primarios, por tanto, se desconocen los grados de asociación y sus efectos directos e indirectos para las características biométricas del fruto y semilla para esta especie de actual potencial agroforestal e importancia en la industria farmacológica, medicinal y alimenticia (Gupta et al., 2019a, 2019b; Chomicki et al., 2020). Por lo que, la búsqueda de alternativas de mejoramiento genético que faciliten o sirvan de base para contribuir a los estudios de esta área de investigación puede ser de gran ayuda en las comunidades de aprovechamiento de esta especie. Por lo anterior, el presente trabajo tuvo como objetivo estimar las correlaciones entre 14 características biométricas del fruto y semilla, al igual que los efectos causales directos e indirectos de las variables estudiadas con respecto al peso del fruto en B. hispida, para contribuir con información básica para la selección y mejoramiento genético de la especie.
MATERIALES Y MÉTODOS
Localización y duración. El estudio se realizó entre agosto de 2016 y julio de 2017, en lotes experimentales y el Laboratorio de Genética Vegetal y Fitomejoramiento de la Universidad de Córdoba (Montería, Colombia), ubicada en la zona media del valle del Sinú, a 8°52’ de latitud norte y 76°48’ longitud oeste, a una altura de 13 msnm. La zona ecológica corresponde al bosque seco tropical con temperatura promedio de 28ºC, humedad relativa del 84% y precipitación anual de 1200 mm (Palencia et al., 2006).
Material genético. La investigación se realizó mediante la evaluación agronómica de 10 familias de hermanos medios (FHM), con cinco repeticiones. Las 10 FHM se originaron de 10 plantas seleccionadas por su valor fenotípico de un lote comercial en 2016.
Variables de respuestas. Se evaluaron 14 características biométricas, ocho del fruto y seis de la semilla de B. hispida. En cada unidad experimental se tomaron al azar cinco frutos de libre polinización, a los cuales se les midieron ocho rasgos biométricos: largo del fruto (LAF), ancho del fruto (ANF), grosor del fruto (GRF), grosor de la pulpa (GPF) y cavidad semilla (CSF), con la ayuda de reglas y cintas métricas a escala de centímetros y milímetros. Se utilizó una balanza marca BJ 410C, para estimar el peso del fruto (PEF), peso de semillas (PSF) y número de semillas (NSF).
De cada fruto cosechado en cada parcela fueron seleccionadas 50 semillas para formar un compuesto balanceado a las cuales se les midieron seis variables biométricas: ancho de semilla (AS), largo de semilla (LS) y grosor de semilla (GS), empleado un nonio Spictools®. Asimismo, se estimó el peso de una semilla (P1S) y el volumen de una semilla (V1S). Esta última variable se midió como el incremento promedio en volumen que se genera en una probeta con volumen de agua destilada conocido, cuando se introdujo una muestra de 100 semillas tomadas al azar de cada parcela de cada FHM. La densidad de una semilla (D1S) fue estimada a partir de la relación (P1S/V1S).
Diseño experimental y análisis de datos. Para la evaluación de características biométricas de las FHM se utilizó el diseño completamente aleatorizado con cinco repeticiones. El tamaño de cada parcela fue de tres surcos de ocho metros de largo, con distancias de siembra de 2 m por 2 m en cuadrado.
Con los datos de las 14 variables de respuesta, se realizaron análisis de varianza para un modelo de efectos fijos, al igual que estimaciones de los coeficientes de las correlaciones fenotípicas, genotípicas y el análisis de sendero. Se utilizó el programa computacional de libre acceso GENES versión Windows GENES V.2016.6.0 (Cruz, 2016). Las estimaciones de las correlaciones fenotípicas y genéticas están dadas por las Ecuaciones 1 y 2:

En donde: r(xy) y COV (xy) son las correlaciones y covarianzas fenotípicas y genéticas entre los caracteres X e Y, respectivamente; "σ2"(x) y "σ2" (y) son las varianzas fenotípicas y genéticas de X e Y, respectivamente.
Se realizó un análisis de sendero para el sistema: PEF como variable efecto (Y) en función de las variables causas: LAF (X1), ANF (X2), GRF (X3), GPF (X4), CSF (X5), PSF (X6) y NSF (X7). El análisis de sendero se originó a partir del uso de las matrices de correlaciones fenotípicas entre tales variables. Para estimar los efectos directos (coeficientes de sendero: Pi) en el análisis de sendero, GENES utiliza la matriz de correlación fenotípica, la descompone y la organiza en el siguiente sistema de matrices (Ecuación 3) (Cruz, 2016):
P = A-1R (Ec. 3)
Dónde: A-1 es la inversa de la matriz de correlaciones (entre cada una de las variables causas), R es el vector de coeficientes de correlaciones entre las variables causas con la variable efecto, y P es el vector coeficientes de sendero.
La descomposición de los coeficientes de correlación de cada una de las variables causas con la variable efecto (rXiY), en sus componentes: efecto directo (Pi) y el efecto indirecto (Ei), permite mediante el despeje de las Ecuaciones 4, 5, 6, 7, 8, 9 y 10, estimar los respectivos efectos indirectos de cada variable causa (Ei):causas con la variable efecto (rXiY), en sus componentes: efecto directo (Pi) y el efecto indirecto (Ei), permite mediante el despeje de las Ecuaciones 4, 5, 6, 7, 8, 9 y 10, estimar los respectivos efectos indirectos de cada variable causa (Ei):
rx1y = P1 + E1: para con LAF (Ec. 4)
rx2y = P2 + E2: para con ANF (Ec. 5)
rx3y = P3 + E3: para con GRF (Ec. 6)
rx4y = P4 + E4: para con GPF (Ec. 7)
rx5y = P5 + E5: para con CSF (Ec. 8)
rx6y = P6 + E6: para con PSF (Ec. 9)
rx7y = P7 + E7: para con NSF (Ec. 10)
El coeficiente de sendero debido a los efectos residuales o a otras variables no consideradas en el estudio (h), se estimó mediante la Ecuación 11:
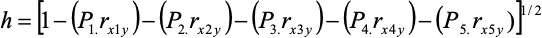
RESULTADOS Y DISCUSIÓN
Análisis de varianza y valores promedio
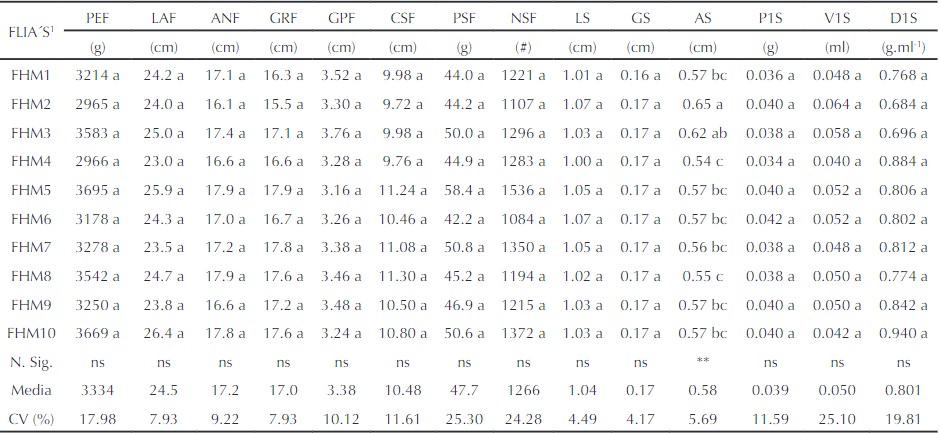
El análisis de varianza para las 10 familias de hermanos medios (FHM) y las 14 variables consideradas en el estudio, se presenta en la Tabla 1. Se detectaron diferencias altamente significativas (p < 0,01) entre familias FHM sólo para AS. Ello indica que al menos una de las FHM presentó promedios estadísticamente diferentes para el AS, más no para las otras variables.
Los valores promedio para las 14 características biométricas del fruto y la semilla son muy similares a los reportados por Tadkal et al. (2019) y Pradhan et al. (2020b). Para AS, la FHM2 difirió significativamente del resto, excepto de FHM3. En las demás variables no se encontraron diferencias significativas entre FHM, situación que refleja en general escasa variabilidad y, por consiguiente, alta homogeneidad fenotípica.
Resultados contrarios se han reportado en la misma especie en algunos estudios (Tadkal et al., 2019; Pradhan et al., 2020b), con diferencias significativas entre genotipos para la mayoría de los caracteres considerados del fruto y de las semillas. Esta poca variabilidad genética reduce la posibilidad de avance genético mediante la selección de familias, especialmente en el (PEF), (GPF) y (CSF), los cuales son de mayor interés comercial en B. hispida (Pradhan et al., 2018; Tadkal et al., 2019). Esta situación posiblemente obedece a que los frutos que originaron las FHM se originaron de polinización cruzada entre pocas plantas o de una sola procedencia, lo que se conoce como efecto fundador (Matute, 2013).
Por lo anterior, el aumento de la variabilidad genética por medio de la introgresión de genes puede constituir el complemento ideal de un programa de mejoramiento genético en B. hispida en el Caribe húmedo colombiano, cuando el objetivo sea aumentar los caracteres biométricos de los frutos y semillas de forma sostenida (Lovely et al., 2017; Pradhan et al., 2018).
Correlaciones
Los coeficientes de correlación fenotípica entre los caracteres biométricos del fruto y la semilla, se presentan en la Tabla 2. Se detectaron correlaciones fenotípicas de diferente magnitud, sentido y significancia estadística entre los caracteres estudiados. De las 91 correlaciones fenotípicas posibles, sólo 19 (20,9%), resultaron significativas al 1% o 5% de probabilidad, lo cual obedece a factores ambientales y genéticos, sobre el grado de asociación entre los pares de variables estudiadas (Hallauer et al., 1988; Falconer y Mackay, 1996).
El mayor número de correlaciones fenotípicas significativas y directas, se presentaron entre las características biométricas del fruto (13/19= 68,4%), con valores entre 0,64* (PEF con NSF) y 0,94** (PSF con NSF), mientras que el grado de asociación entre las características de la semilla, presentaron el menor número de correlaciones significativas (6/19 = 31,6%), con sentido directo (0,66* a 0,88**) e inverso (-0,73* a -0,89**). Es de resaltar, que no hubo asociación significativa entre las características del fruto y de la semilla, lo cual permite inferir que, en las poblaciones evaluadas, los dos grupos de variables presentaron control genético y efecto ambiental independiente, como lo reportan Hang-Vu et al. (2019).
En el fruto, se presentaron correlaciones fenotípicas significativas directas entre: PEF y NSF, CSF, PSF, GRF, LAF y ANF, con coeficientes que oscilaron entre 0,64* (NSF) y 0,91** (ANF); LAF y ANF con grados de asociación de 0,73*; ANF y CSF (0,76*) y GRF (0,81**); GRF y NSF, PSF y CSF, con coeficientes de correlación entre 0,71* y 0,87**; PSF y NSF (0,94**). Los anteriores resultados concuerdan parcialmente con los reportados en otras especies (Balaguera-López et al., 2020; Pradhan et al., 2020b). No obstante, es necesario tomar estos resultados con cierta reserva, ya que según lo manifestado por Falconer y Mackay (1996) y Hallauer et al. (1988), las correlaciones fenotípicas tienen poco valor práctico y pueden conllevar a errores, por ser el resultado de efectos tanto de naturaleza genética como ambiental.
| VAR1 | LAF | ANF | GRF | GPF | CSF | PSF | NSF | LS | GS | AS | P1S | V1S | D1S |
| PEF | 0.86** | 0.91** | 0.78** | 0.11 | 0.67* | 0.71* | 0.64* | -0.02 | 0.44 | -0.17 | 0.28 | -0.08 | 0.15 |
| LAF | 0.73* | 0.46 | -0.16 | 0.45 | 0.59 | 0.50 | 0.15 | 0.41 | 0.06 | 0.45 | 0.02 | 0.16 | |
| ANF | 0.81** | -0.01 | 0.76* | 0.58 | 0.60 | -0.16 | 0.25 | -0.43 | 0.10 | -0.29 | 0.23 | ||
| GRF | -0.07 | 0.87** | 0.71* | 0.71* | -0.08 | 0.50 | -0.58 | 0.13 | -0.42 | 0.45 | |||
| GPF | -0.28 | -0.19 | -0.22 | -0.30 | -0.15 | 0.26 | -0.29 | 0.28 | -0.50 | ||||
| CSF | 0.55 | 0.48 | 0.15 | 0.36 | -0.51 | 0.35 | -0.23 | 0.31 | |||||
| PSF | 0.94** | 0.16 | 0.41 | -0.09 | 0.15 | -0.05 | 0.17 | ||||||
| NSF | -0.14 | 0.25 | -0.32 | -0.13 | -0.35 | 0.38 | |||||||
| LS | 0.49 | 0.55 | 0.83** | 0.66* | -0.35 | ||||||||
| GS | -0.01 | 0,70* | -0.01 | 0.34 | |||||||||
| AS | 0,32 | 0.88** | -0.73* | ||||||||||
| P1S | 045 | -0.08 | |||||||||||
| V1S | -0.89** |
En la semilla, los grados de asociación significativos y positivos fueron: LS con V1S y P1S (0,66* y 0,83**, respectivamente), y AS con V1S (0,88**). De igual manera, las correlaciones significativas con signo negativo fueron: D1S con AS y V1S (-0,73* y -0,89**, respectivamente). Estos resultados son similares a los reportados en otros estudios (Hang-Vu et al., 2019; Balaguera-López et al., 2020).
La ausencia de significancia en la correlación entre el PEF y GPF (0,11), no es muy común, dado que el GPF al ser uno de los componentes primarios importantes del PEF, se esperaba que resultara con correlación fenotípica alta, directa y significativa; esto posiblemente obedezca a efectos enmascaradores indirectos que tienen otras variables, sobre el real nivel de asociación entre ellas, como lo explican Hang-Vu et al. (2019).
Las magnitudes, sentido y significancia de las correlaciones entre el PEF y LAF, ANF, GRF, CSF, PSF y NSF (rF≥0,64) son de gran importancia en los procesos de selección, manejo y conservación de las semillas. Para el fitomejorador y el productor, la selección de genotipos de alta producción de semillas y pulpa representan componentes valiosos para el consumo y transformación (Chomicki et al., 2020; Pradhan et al., 2020a), y como garantía de supervivencia de la especie (Nansikombi et al., 2019).
En la producción de híbridos F1 o variedades, es posible realizar indirectamente la selección de plantas con altos PSF Y NSF, a través de PEF y GRF, las cuales son mucho más fáciles de medir, comparadas con PSF y NSF, que requieren de mayor tiempo, son más difíciles y costosas de estimar; esto es importante cuando el órgano de interés es la semilla para su consumo cuando son procesadas, en la agroindustria o en la producción de plántulas en viveros (Balaguera- López et al., 2020; Chomicki et al., 2020). En este sentido, Espitia-Camacho et al. (2021), reportan bajos porcentajes de germinación y problemas de dormancia, que requieren tratamientos pregerminativos para poder lograr una alta germinación.
El grado de asociación significativa y directa del V1S con LS y AS, y la LS con P1S, es de gran interés, ya que el peso y el volumen de una semilla, por lo general están asociados con un mayor porcentaje de germinación y vigor de las plántulas, en razón a que semillas grandes y pesadas contienen mayores cantidades de carbohidratos en el endospermo o cotiledones, que reflejan la disponibilidad mayor fuente de energía para estimular la germinación, emergencia, vigor, supervivencia y crecimiento de las plántulas (Balaguera-López et al., 2020; Chomicki et al., 2020). Estas correlaciones han sido detectadas en algunos trabajos (Balaguera-López et al., 2020; Chomicki et al., 2020).
Considerando lo anterior, Hallauer et al. (1988), señalan que la diferencia entre las correlaciones para un mismo grupo de variables, aun en una misma especie, es común y se explican por una influencia moderada o efecto de enmascaramiento de unas variables sobre otras, interacción de factores ambientales y/o factores genéticos no aditivos (interacciones genéticas), los cuales subestiman o sobreestiman la verdadera expresión del nivel de correlación entre las variables estudiadas, ya que esos factores ambientales pueden afectar a las variables en estudio a través de un mismo o diferentes mecanismos fisiológicos, lo cual justifica aplicar el análisis de sendero en el estudio de las correlaciones.
Respecto a las correlaciones genéticas, al presentarse estimaciones de varianzas genéticas (VG) con valores negativos para varios caracteres como ANF, PESF y NSF, no se estimaron correlaciones genéticas válidas. El origen de tales varianzas negativas se explica por tres posibles causas: i) poca variabilidad genética y alta homogeneidad fenotípica en las 10 FHM evaluadas (Tabla 1), ii) influencia del tamaño de muestra en la estimación de las variables y iii) la estimación de VG en la Ecuación VG = VF - VA, es decir, al restar una varianza ambiental (VA) de mayor magnitud que la varianza fenotípica (VF) resulta una VG negativa en virtud del modelo de componentes de varianza comúnmente usado en este tipo de estimaciones.
Según Hallauer et al. (1988), las estimaciones de varianzas negativas son muy comunes en estudios de varianzas y componentes de varianzas en plantas, sugiriendo a su vez que las estimaciones de valores negativos puedan ser reportados como cero o negativos. Por ello sugiere que ambas valoraciones son válidas, teniendo en cuenta que estos casos ocurren y que el investigador no debe asumir que ha realizado algún cálculo errado o que los resultados experimentales carezcan de validez. Esta situación condujo también a que el análisis de sendero se hiciera sólo con base en las correlaciones fenotípicas, tal como ha sucede en otros estudios (Tratenmuller et al., 2019; Hang-Vu et al., 2019).
Análisis de sendero para el peso del fruto (PEF)
La descomposición de las correlaciones fenotípicas para el PEF con las otras variables del fruto de B. hispida, en sus efectos directos e indirectos, se muestran en la Tabla 3. Para la variable efecto PEF, se realizaron varios modelos en los que se incluyeron progresivamente un número mayor de variables causas (componentes primarios y secundarios del PEF), las estimaciones de los coeficientes de determinación (R2) y los efectos residuales (h) para cada modelo. Esto condujo a la decisión de incluir sólo en el análisis las variables LAF, ANF, GRF, GPF, CSF, PSF y NSF, como variables causas o explicativas. El R2 para el análisis de sendero con base en las correlaciones fenotípicas fue del 99%, lo cual es idóneo para un modelo no muy complejo e indica a la vez la importancia de los componentes primarios considerados en la definición del PEF.
| Variables | Correlaciones fenotípicas | rF | ||||||
| LAF | ANF | GRF | GPF | CSF | PSF | NSF | con PEF | |
| LAF | 0.52 | 0.18 | 0.14 | -0.04 | 0.00 | 0.13 | -0.07 | 0.86** |
| ANF | 0.38 | 0.24 | 0.25 | 0.00 | -0.01 | 0.13 | -0.08 | 0.91** |
| GRF | 0.24 | 0.20 | 0.31 | -0.02 | -0.01 | 0.15 | -0.09 | 0.78** |
| GPF | -0.08 | 0.00 | -0.02 | 0.22 | 0.00 | -0.04 | 0.03 | 0.10 |
| CSF | 0.24 | 0.19 | 0.27 | -0.06 | -0.01 | 0.12 | -0.06 | 0.67* |
| PSF | 0.31 | 0.14 | 0.22 | -0.04 | -0.01 | 0.22 | -0.12 | 0.71* |
| NSF | 0.26 | 0.15 | 0.22 | -0.05 | 0.00 | 0.20 | -0.13 | 0.64* |
| R2 = 0.99 | h= 0.06 | |||||||
Se evidenció mayor importancia de los efectos indirectos sobre los efectos directos en las variables explicativas, dado que los efectos indirectos explican el nivel de correlación significativa en cuatro de las variables: ANF, CSF, PSF y NSF, mientras que los efectos directos lo hacen únicamente en dos caracteres LAF y GRF. El grado y sentido de correlación entre PEF y LAF a nivel fenotípico (0,86**), es explicado vía efecto directo (0,52) en un 60,5%. Situación diferente se observa para la asociación entre PEF y ANF (0,91**), la cual es explicada por el efecto directo (0,24) en un 26,4% y por los efectos indirectos vía LAF (0,38) y GRF (0,25) en un 69,2%. Resultado similar se observa en la asociación entre PEF y GRF (0,78**), la cual es explicada por el efecto directo (0,31) en un 39,7% y por los efectos indirectos vía LAF (0,24) y ANF (0,20) que juntas explican tal asociación en 56,4%.
La correlación de las variables PEF y PSF (0,71*), a través de los efectos indirectos vía LAF (0,31) y GRF (0,22) explican un 74,6% el nivel de asociación, mientras que el efecto directo (0,22) apenas el 31%. Las correlaciones de PEF con CSF y NSF (0,67* y 0,64*, respectivamente), se explican principalmente por los efectos indirectos vía LAF (0,24 y 0,26, respectivamente) y GRF (0,27 y 0,22, respectivamente), los cuales representan en total el 75% o más del nivel de asociación. Resultados parcialmente similares han sido reportados en B. hispida (Pradhan et al., 2020b) y en otras especies (Tratenmuller et al., 2019; Hang-Vu et al., 2019).
Desde el punto de vista de las variables causas, el LAF exhibió los mayores efectos directos e indirectos sobre el PEF, en comparación con las demás variables causas en el análisis de sendero (Tabla 3); los cuales son coherentes con los reportados en B. hispida (Pradhan et al., 2020b) y en algunas hortalizas, leguminosas y frutales (Hang-Vu et al., 2019).
CONCLUSIONES
En la población estudiada de B. hispida, los caracteres biométricos del fruto y semilla presentaron poca variabilidad genética.
El largo y peso del fruto puede ser utilizado como criterio de selección, para la obtención de genotipos de mejor producción, cavidad de semillas, peso de semillas y número de semillas por fruto.
El LAF exhibió los mayores efectos directos e indirectos sobre el PEF, en comparación con las demás variables causas en el análisis de sendero, por lo que se puede inferir que el PEF y LAF pueden ser usados como criterios de selección, en el mejoramiento genético de B. hispida para la selección y obtención de genotipos con mayor producción y CSF, PSF y NSF.
REFERENCIAS
Balaguera-López, H.E., Fischer, G., y Magnitskiy, S. 2020. Seed-fruit relationships in fleshy fruit species: Role of hormones. A review. Revista Colombiana de Ciencias Hortícolas. 14(1):90-103. https://doi.org/10.17584/rcch.2020v14i1.10921
Chomicki, G.; Schaefer, H., Renner, S. 2020. Origin and domestication of Cucurbitaceae crops: insights from phylogenies, genomics and archaeology. New Phytologist. 226(5):1240-1255. https://doi.org/10.1111/nph.16015
Cruz, C.D. 2016. Programa Genes V.2014.6.1 - Aplicativo computacional em genética e estatística. Disponible desde internet en http://www.ufv.br/dbg/genes/genes.htm
Ekeke, C., Ogazie, C.A., Agbagwa, I.O. 2019. Anatomical and Phytochemical Studies on Benincasa hispida (Thunb.) Cogn. (Cucurbitaceae). NotulaeScientia Biologicae. 11(1):102-111. https://doi.org/10.15835/nsb11110394.
Espitia-Camacho, M., Villalba-Soto, A., Araméndiz-Tatis, H., Cardona-Ayala, C. 2021. Morphometry, viability and germination of seeds of Benincasa hispida [(Thunb.) Cogn.]. Revista Colombiana de Ciencias Horticolas. 15(1):21-37. https://doi.org/10.17584/rcch.2021v15i1.11724.
Falconer, D.S., Mackay, T. 1996. Introduction to Quantitative Genetics. Fourth edition. Prentice Hall (USA). 464p.
Gupta, P., Chikkala, S., Kundu, P. 2019a. Ash gourd and its applications in the food, pharmacological and biomedical industries. International Journal of Vegetable Science. 27(1):44-53. https://doi.org/10.1080/19315260.2019. 1699222.
Gupta, S., Patel, R., Bhave, M. 2019b. Development of A nutraceutical product using Benincasa hispida. International Journal of Advance Research, Ideas and Innovations in Technology. 5(1):579-582.
Hallauer, A.R., Carena, M.G., Miranda, F.J.B. 1988. Quantitative Genetics in Maize Breeding. Second edition. Iowa State Univ. Press Iowa. p169-184.
Hang-Vu, T.T., Cham-Le, T.T., Hoa-Vu, D., Nguyen, T.T., Pham, T.G. 2019. Correlations and Path Coefficients for Yield Related Traits in Soybean Progenies. Asian J Crop Sci. 11(1):32-39. h t t p s : / / d x . d o i . o r g / 1 0 . 3 9 2 3 / pjbs.2020.425.438.
Lovely, B., Vijayaraghava, K. 2017. Estimates of genetic variability, heritability and genetic advance for yield and yield component traits in ash gourd [Benincasa hispida (Cogn.)] genotypes. Agricultural Science Digest. 37(4):270-274. DOI: 10.18805/ag.D-4676.
Matute, D. R. 2013. The role of founder effects on the evolution of reproductive isolation. Journal of evolutionary biology. 26(11):2299-2311. https://doi.org/10.1111/jeb.12246.
Nagaraju, K., Saraswati, T., Sharma, R., Bommesh, J.C. 2016. Correlation and path analyses among scented and non- scented ash gourd [Benincasa hispida (Thunb.) Cogn.] genotypes. International Journal of Agricultural Sciences. 8(61):3494-3498. Available online at http://www.bioinfopublication.org/ jouarchive.php?opt=&jouid=BPJ0000217
Nansikombi, N., Muyonga, J.H., Byaruhanga, Y.B. 2019. Association between Fruit Characteristics and Postharvest Stability of Different Pumpkin (Cucurbita) Species. Journal of Food Research. 8(4):131-145. https://doi.org10.5539/jfr.v8n4p131
Palencia, G., Mercado, T., Combatt, E. 2006. Estudio agroclimático del departamento de Córdoba. Editorial Caribe.126p.
Pradhan, K., Nandi, A., Rout, S., Tripathy, B. 2020a. Ash gourd - an under exploited potential crop. Dogo Rangsang Research Journal. 10(06):142-151.
Pradhan, K., Nandi, A., Das, S., Sahu, G.S., Rout, S. 2020b. Correlation and Path Analysis of the Yield Contributing Characters of Different Ash Gourd [(Benincasa hispida (Thunb.) Cogn.] Germplasm. Ind. J. Pure App. Biosci. 8(3):248-254. http://dx.doi.org/10.18782/2582- 2845.8130
Pradhan, K., Nandi, A., Das, S., Sahu, G.S., Sarkar, S., Patnaik, A. 2018. Genetic variability and varietal performance in Ash Gourd [Benincasa hispida (Thunb) Cogn.] genotypes. The Bioscan. 13(2):791-794.
Tadkal, R., Beaulah, A., Krishnamoorthy, V., Thangaraj, K. 2019. Evaluation of ash gourd (Benincasa hispida) (Thunb.) (Cogn.) genotypes for growth and yield under pandal system of cultivation. International Journal of Chemical Studies. 7(3):2933-2937.
Tratenmuller, J.W., Péllico-Netto, S., Balbinot, R., Corte, A.P.D., Borella, J. 2019. Path analysis applied to evaluation of biomass estimates in subtropical forests of Brazil. Floresta. 49(3):587-596. http://dx:doi.org/10.5380(rf.v49i3.60782
Información adicional
Conflicto de intereses: El manuscrito fue preparado y revisado con la
participación de todos los autores, quienes declaramos que no existe conflicto de intereses que ponga en riesgo la validez de los resultados presentados.

